自然界中的一些微生物可以通过合成纳米导线与环境介质及其它微生物进行长距离电子传递及种间电子传递。但是,目前已发现的可以合成纳米导线进行长距离电子传递的细菌资源极其缺乏,且均为革兰氏阴性菌,而对于环境中广泛存在且发挥重要作用的革兰氏阳性菌是否具有这一功能一直没有答案。针对此关键问题,广东省科学院微生物研究所研究员杨永刚联合丹麦、比利时及国内多个研究团队于2021年在Nature Communications上发表题为“Long-distance electron transfer in a filamentous Gram-positive bacterium”研究论文。
第一部分:内容解读
摘要:在自然和工程过程中,革兰氏阴性细菌中可以观察到长距离细胞外电子传递。这种电子传递可以由导电蛋白附属物介导(简称单细胞细菌,如地杆菌Geobacter)或通过导电细胞包膜(在丝状多细胞电缆细菌中)。本研究表明,长线形革兰氏阳性菌Lysinibacillus varians GY32,一种丝状单细胞革兰氏阳性细菌,能够进行双向细胞外电子转移。在微生物燃料电池中,L. varians可以形成厘米范围的导电细胞网络,当它生长在石墨电极上时,细胞长度可以达到1.08毫米。原子力显微镜和微电极分析表明,电导率与皮脂样蛋白质附属物有关。本研究结果表明,长距离电子转移不仅限于革兰氏阴性菌。
研究背景:电子传递对于能量产生和生命新陈代谢至关重要。细菌在将电子转移到各种化学物质和保存能量的能力方面具有多功能性。除了用细胞内化学物质(如氧气、硫酸盐或硝酸盐)呼吸外,细菌还可以用外部接触的化学物质(如矿物颗粒)进行呼吸(即胞外电子传递,简称EET)。一些单细胞细菌能够以数十微米的距离向化学物质或其他微生物细胞进行长距离电子转移(简称LDET),多细胞的电缆细胞则可以在厘米范围内传输电子。
目前已经确定了两种细菌长距离电子传递的方式:由相对较短的单细胞细菌(如地杆菌Geobacter)和由丝状多细胞的电缆细菌形成的导电包膜产生的蛋白质纳米导线导电。由Geobacter和其他几种微生物产生的导电蛋白纳米导线可以在数十微米距离内转移电子。这些蛋白质纳米导线可以形成导电网络,并有助于由不同微生物组成的生物膜或聚集体中的细胞间电子直接转移。电缆细菌通常形成厘米长度的细丝,它们由数千个细胞首尾相连,进而可以耦合厌氧沉积物中的硫化物氧化和水生系统中沉积物表面的氧气还原。电缆细菌的周质纤维已被证明可以导电,并且可以促进LDET。细菌LDET的广泛发生影响着地球上自然和工程环境中的微生物群落和生物地球化学过程。到目前为止,能够进行LDET的细菌都被报道是革兰氏阴性的,尽管革兰氏阳性细菌无处不在,有些能够进行EET,由于革兰氏阳性菌具有不同的细胞表层,它们没有被认为可以形成LDET。
本文发现了长线形革兰氏阳性菌GY32发生的额外的细菌LDET方式,该方式能够进行双向EET。在微生物燃料电池(简称MFC)中,菌株GY32可以形成厘米范围的导电细胞网络,该网络由具有导电纳米导线状附属物的绝缘丝状细胞组成。
方法部分:本研究主要使用电化学手段设置生物燃料电池和生物阴极电化学系统评估沉积物中GY32菌株的电子传递过程,同时用电镜细菌形貌变化分析电子传递发生部位,利用转录分析GY32菌株上发生电子传递的主要成分。
对于含有液体介质的MFC,设置了一个双壳室微生物燃料电池MFC装置(装置图1上)。其中,阳极和阴极采用石墨板。阳极培养基中利用5 mM的甲酸盐或乙酸盐作为唯一的电子供体,厌氧条件下接种GY32菌株。阴极电解质中为50 mM铁氰化钾的PBS试剂。MFC运行期间通过测定阳极培养物的总蛋白质浓度来确定阳极细菌生长,利用高效液相色谱系统定期评估阳极里电子供体消耗。在上述MFC基础上,为了测试GY32对极化电极的EET能力,用电化学工作站替换外部电阻,并在每个阳极室中放置Ag/AgCl参比电极。阳极在0.4 V(VS SHE)下极化,并持续记录阴阳极之间电流大小。此外也测量了工作电极上GY32生物膜的循环伏安曲线。除了MFC装置,本研究还设置了一个双室生物阴极电化学系统BCES装置(装置图2下)。阴极中添加2 mM柠檬酸铁进行Fe(III)还原,并在厌氧条件下接种GY32菌株。通过使用电化学工作站控制阴极电位恒定为−0.6 V(vs SHE)。
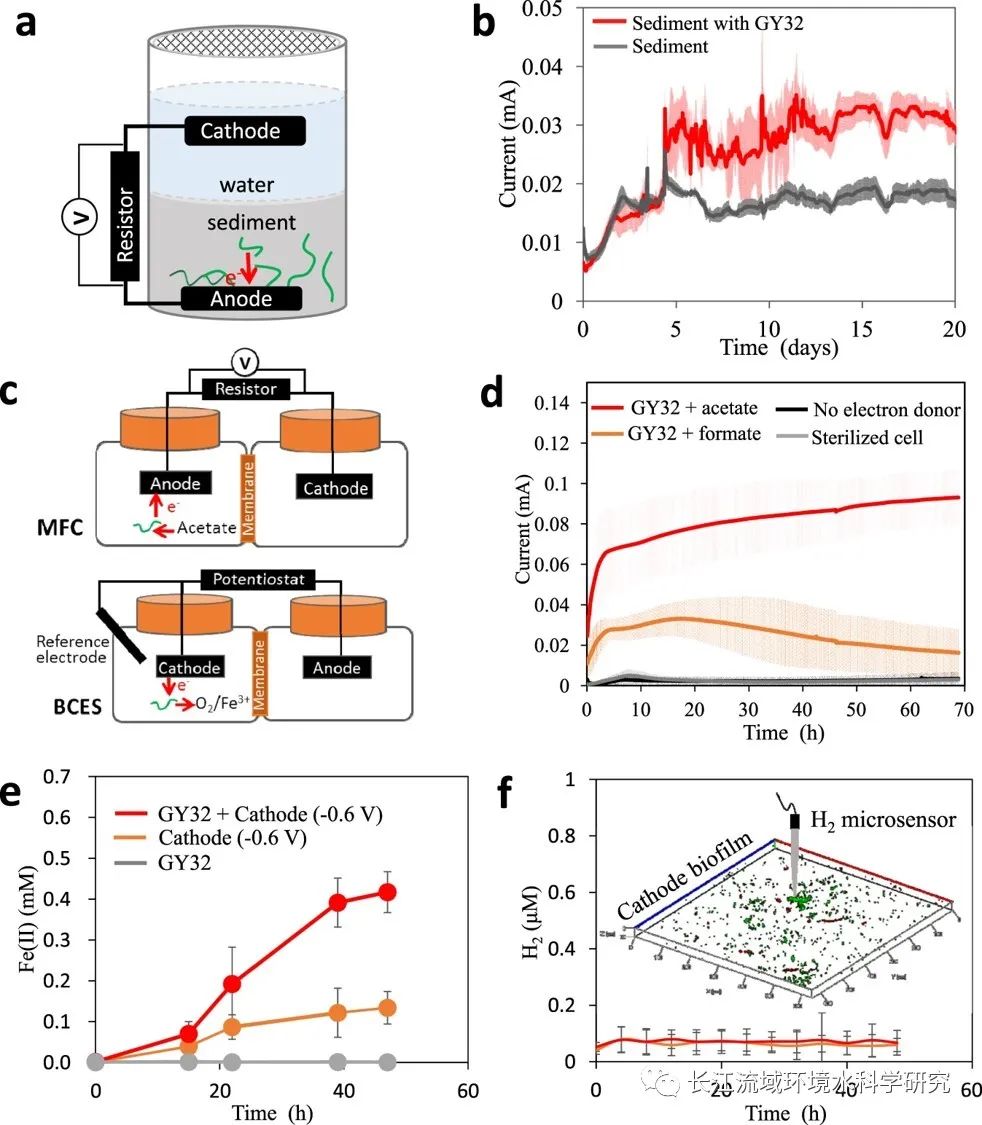
装置图2C.在含有液体介质的MFC(上层反应器)和BCES(下层反应器)中GY32胞外电子传递(EET)示意图
沉积物生物燃料电池(简称SMFCs)装置如图2A所示。灭菌或未灭菌的沉积物中预先接种GY32菌株,普通石墨毡用于沉积物中的阳极和水中的阴极。为了排除环境微生物,烧杯上覆盖着八层纱布。使用电阻为1000 Ω的钛线来连接所有SMFC中的阳极和阴极。通过电阻器的电流由万用表监控。
结果:
L. varians GY32可以形成极长的细胞
菌株GY32是从淡水沉积物中分离出来的单细胞丝状革兰氏阳性细菌。前人报道GY32细胞长度达到约500 μm,直径均匀(∼0.5 μm)。本研究发现在厌氧条件下石墨电极作为MFC中唯一的电子受体时,GY32菌株可以生长为极长的细胞,最长的GY32细胞为1.08 mm(Fig.1a)且呈现单细胞结构(Fig. 1b, c)。GY32的菌丝保持 8 μm−1 的表面体积比,类似于棒状细菌。更高的表面体积比可以支持单位细胞体积内更快的养分和废物交换速度,因此细菌生长速度更快。大多数革兰氏阳性细菌细胞壁中具有多层肽聚糖。在GY32中,本研究观察到其表面上具有二维晶体层(Fig. 1d),表明在肽聚糖顶部有保护性S层结构。切片结果(Fig. 1c)表明细胞壁宽度为20-30 nm,可与其他一些革兰氏阳性细菌细胞壁媲美,且比革兰氏阴性细菌(2-7 nm)厚得多。通过透射电子显微镜(TEM)和原子力显微镜(AFM)观察细胞外面细胞壁、毛状或纳米导线状附属物(Fig. 1d-h)发现,这附属物经常结合在一起,并且长度单个附属物可以超过10 μm。附属物的大小和结构与硫还原地杆菌的IV型吡咯烷和多血红素c型细胞色素纳米导线相似。
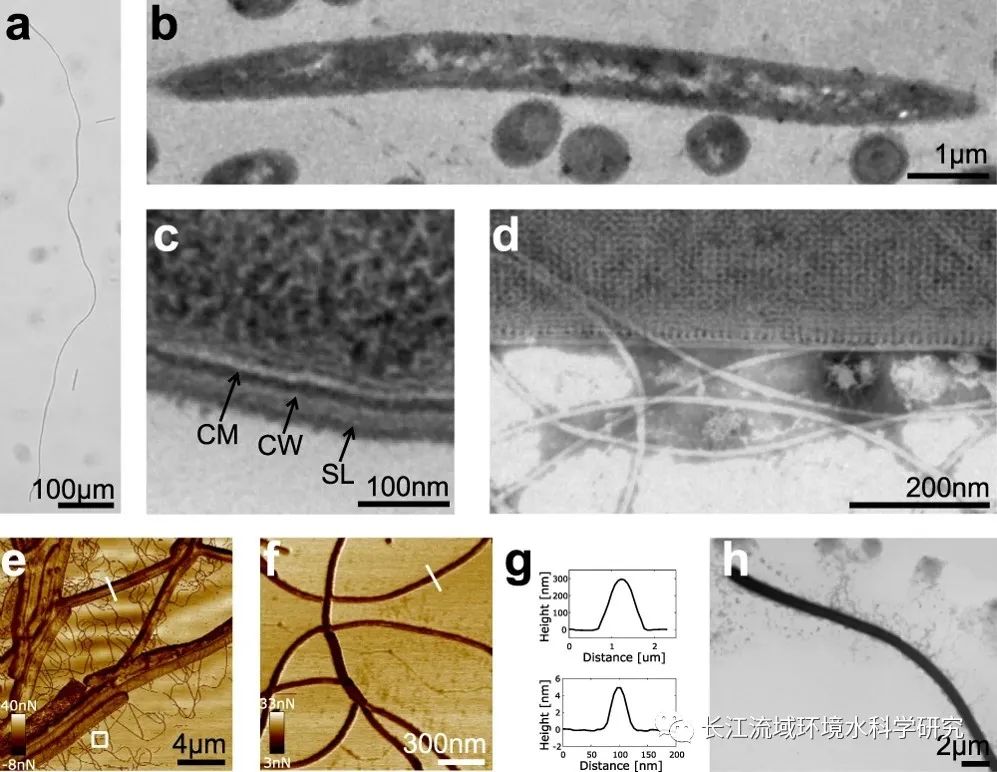
图1 细胞形貌特征
L. varians GY32可以将电子转移到石墨电极上沉积物和液体环境
在含有指定液体介质的MFC中(装置图1),阳极室中浮游细胞的生物量增加与乙酸盐同时消耗和电流的生成表明GY32菌株以石墨阳极作为唯一的电子受体进行呼吸和生长。此外,在乙酸盐作为电子供体的存在下,GY32和MR-1观察到具有较高峰值电流的类似的循环伏安曲线,表明菌株生物膜内产生了氧化还原性物质。本研究进一步测试了在BCES中(装置图2),GY32利用电极作为电子供体柠檬酸铁作为电子受体的能力。相比未极化时,石墨阴极被极化时GY32明显还原Fe(III)(Fig. 2e)。负极化电极中生成的H2被用作电子供体被排除对Fe(III)的还原,这些结果表明,GY32使用阴极作为唯一的电子供体来还原BCES中的Fe(III)。此外, GY32被发现也可以使用极化电极作为电子供体来还原溶解氧。通过与发电能力结合的结果表明,GY32可以作为胞外氧化还原的电子供体子或受体。GY32的这种双向EET能力意味着GY32具有与另一个细菌细胞进行种间电子转移的可能性。
L. varians及其附属物形成导电细胞网络
当将电子转移到MFC中的阳极时, GY32优选在阳极表面周围聚集并形成网络状结构(Fig. 3a)。这些网络状结构可以延伸几厘米从阳极表面进入液体介质。Fig. 3a还显示了纳米导线状的附属物沿着丝状细胞形成子网集群中的不同像元。Geobacter的细菌纳米导线和Shewanella菌株中观察到明显的细胞附着的附属物,表明这些是由蛋白质组成(Fig. 3a)。进一步GY32组成细胞及其附属物被挑选并用于连接两个绝缘预制金电极(Fig. 3b)。当施加0.1 V的固定电压时,形成稳定电流,表明细胞簇具有导电能力。相比之下,对照样品包括(a)热灭活GY32细胞簇,(b)未连接金电极的细胞簇,(c)培养上清液对电压的反应要小得多。导电的GY32细胞簇表明阳极周围的厘米范围细胞网络是导电的。此外使用开尔文探针力显微镜(KPFM)来进一步了解纳米结构的电学性质GY32细胞簇,表明附属物比GY32的包膜导电性强。
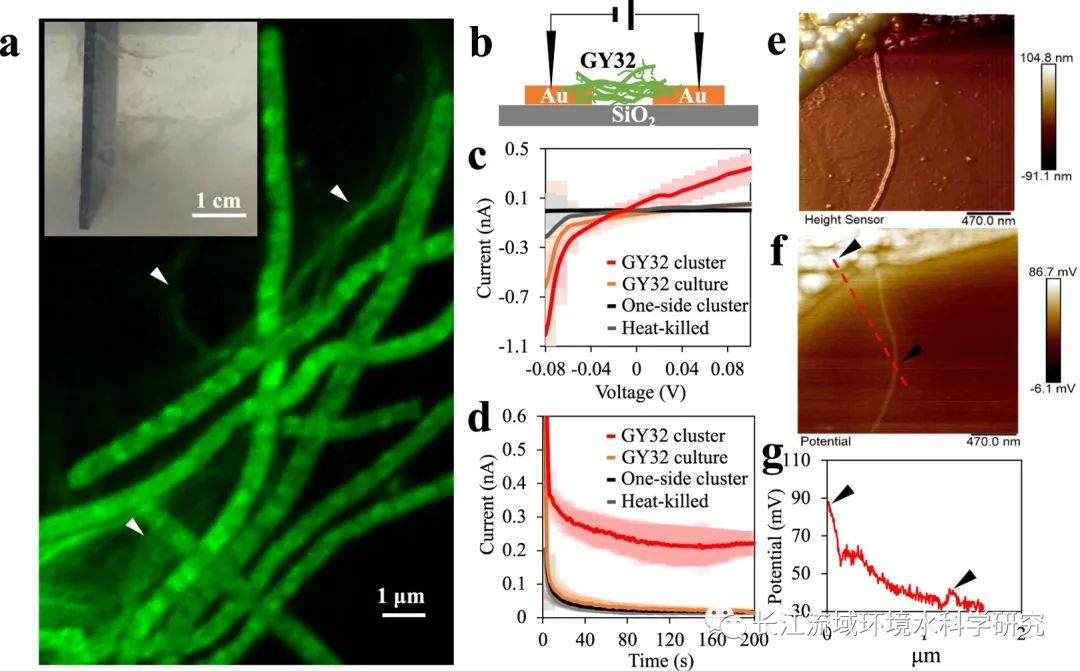
图3. 细丝状和附着在纳米有机上纳米线状细胞GY32网络比较
微电极试验表明GY32附属物的电导率
丝状细胞及其附属物是GY32细胞网络中主要成分。根据以往报道中电缆细菌的导电细胞包膜和地杆菌的导电纳米导线,这两种细胞的电导率被用来评估GY32的包膜和附属物。当使用单个GY32细胞(没有连接电极的附属物)来连接制造的电极阵列时(Fig. 4a,b),在直接电压为0.1 V时没有获得明显的电流。当电压范围为−0.1至0.1 V时(Fig. 4 c),GY32细胞包膜不导电。另一方面,与GY32细胞团簇类似,附属物的电导率在干燥条件下降低。这些结果表明细胞附属物是导电的,可以负责细胞网络的电导率,尽管需要进一步研究来阐明单个附属物的化学成分和电导行为。
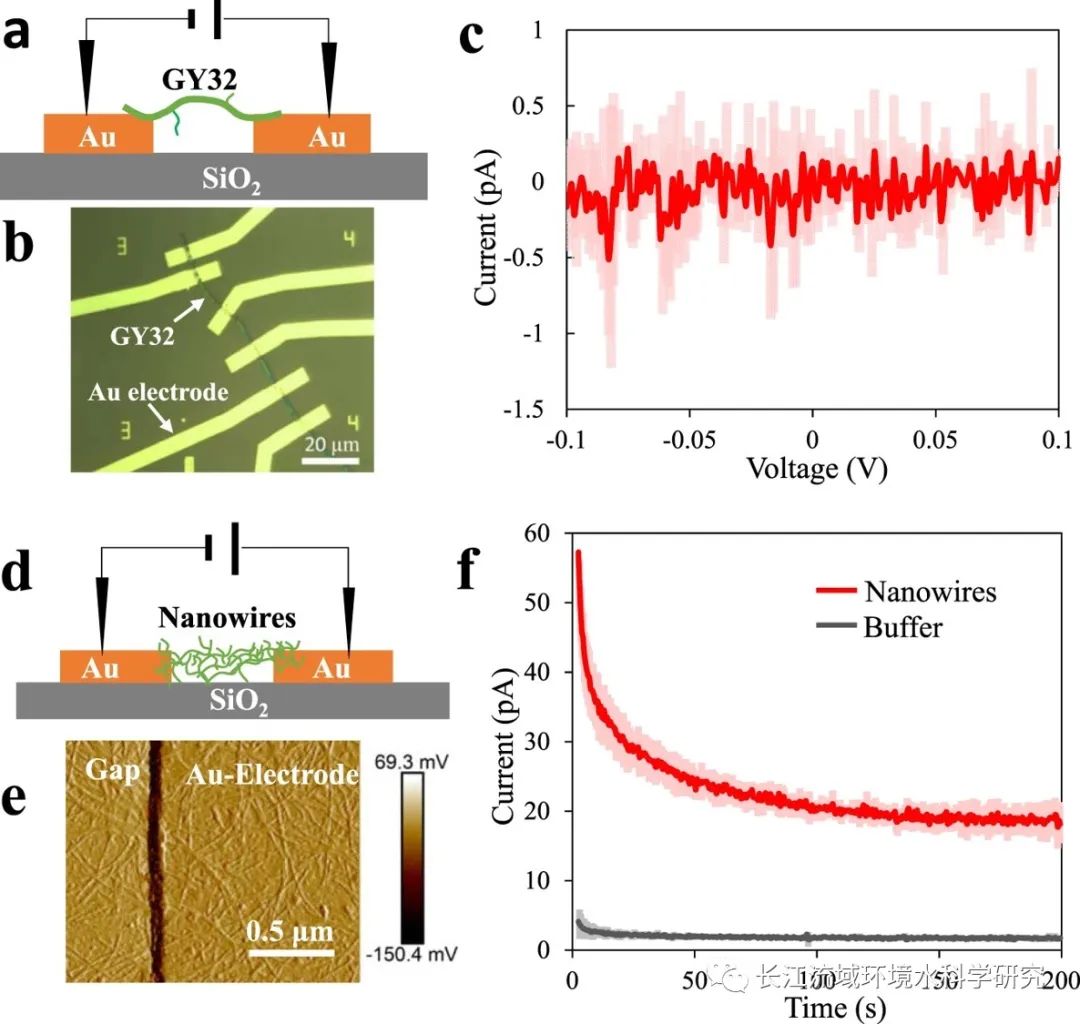
图4 电导测定
参与GY32长距离电子传递的可能成分
细菌可以产生一系列具有明显特征的附属物功能、结构和分子组成,包括鞭毛、纤毛、细胞膜延伸、c型细胞色素聚合物、伪皮林和其他未知纳米导线。EET 方式通常涉及c型细胞色素或电子穿梭,并且已在使用核黄素为电子穿梭体的革兰氏阳性菌中观察到。然而没有研究表明在EET或LDET中涉及革兰氏阳性菌的纳米导线状附属物。由于GY32的纳米导线状附属物太薄而不能成为像MR-1一样的细胞膜延伸,GY32的附属物有一个大小与某些革兰氏阴性细菌(即e-pili)的导电IV型皮毛相似。通过转录分析推测,GY32的导电附属物可能由ComGD组成,并且可能具有与地杆菌的e-pili相似的电导机制。此外,c型细胞色素的线性聚合物可以形成导电附属物。GY32有六种推定的c型细胞色素基因。这些结果表明c型细胞色素可能在GY32的EET中发挥作用。然而,这些c型细胞色素是否以及如何参与在LDET和EET中需要更多的研究。
讨论:
该研究发现,一株分离自电子垃圾污染河流沉积物的长线形革兰氏阳性菌Lysinibacillus varians GY32可以在采用培养液或沉积物构建的生物电化学系统中产出电能。在产电过程中,菌株GY32的细胞进一步伸长(单个细胞长度可超过1 mm),并围绕电极相互缠绕,形成厘米尺度的细胞网络。导电测试发现,菌株GY32聚集形成的细胞网络具有良好的导电特性。通过原子力显微镜和微电极阵列的测试发现,这一导电细胞网络中的细胞是绝缘的,但细胞周身会合成长度可达十几微米的蛋白纳米线,这些蛋白纳米线具有明显的导电性,是GY32细胞网络导电的关键组分。尽管如此,GY32细胞导线的详细组分和导电机理仍有待阐明。
越来越多的证据表明,微生物LDET在自然和工程环境中广泛分布。已经深入研究了两种类型的微生物LDET网络。首先,短细胞(例如,地杆菌和Shewanella)与细胞外纳米导线,这些 LDET 网络在很大程度上依赖于颗粒表面的生物膜形成,并且具有有限的厚度(一般≤0.1 mm)。其次,丝状电缆细菌具有连续连接的细胞和细胞内纳米线。电缆细菌LDET网络可以跨越厘米距离,但主要存在于沉积物-水界面。革兰氏阳性菌GY32的长距离电子传递网络包括丝状单细胞细胞和细胞外纳米线导状附属物,类似于革兰氏阴性细菌两种 LDET 网络模式的组合(Fig. 5)。鉴于GY32是一个兼性厌氧细菌,它可能在氧化和缺氧环境中形成潜在的导电网络。
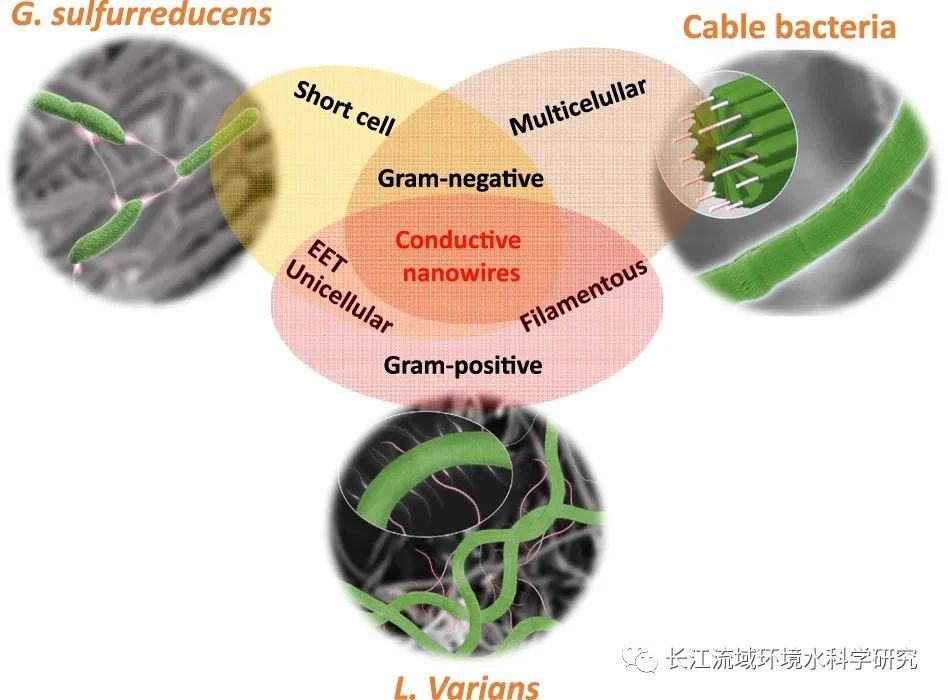
图5 导电网络分析
第二部分:贡献解读
背景问题:自然界中的一些微生物(如电缆细菌cable bacteria和地杆菌Geobacter)可以通过合成纳米导线与环境介质及其它微生物进行长距离电子传递及种间电子传递。这些反应在环境中普遍存在,并且通常相互耦联形成长距离电子传递网络,对元素的生物地球化学循环、污染物降解转化等过程产生重要影响。但是,目前已发现的可以合成纳米导线进行长距离电子传递的细菌资源极其缺乏,且均为革兰氏阴性菌,而对于环境中广泛存在且发挥重要作用的革兰氏阳性菌是否具有这一功能一直没有答案。
核心发现:该研究首次发现了革兰氏阳性细菌通过纳米导线形成长距离电子传递网络,为全面认识自然环境中的微生物长距离电子传递网络提供了重要补充。此外,菌株GY32这种单个细胞长度超过1 mm、且包含多个核区的细胞形态也拓展了人们对细菌形态和分裂机制的认识。
原文来源:Yang Y. G., et al., Long-distance electron transfer in a filamentous Gram-positive bacterium. Nature Communications, 2021, 12(1): 1709.
DOI: 10.1038/s41467-021-21709-z.
本文由中国地质大学(武汉)生物地质与环境地质国家重点实验室袁松虎教授课题组博士生张妍婷解读。受作者能力所限,本文难免有不当之处,敬请各位读者谅解。如疑义、建议或其他方面的学术交流,请与袁松虎教授联系,邮箱yuansonghu622@cug.edu.cn.
